



Hatch Plasticity in Ochlerotatus triseriatus in
Response
to Hatch Stimulation Frequency
Christopher J.
Vitek, Camilo Khatchikian, Todd P. Livdahl
Biology Department,
Clark University, 950 Main St., Worcester, MA 01610
Introduction
Phenotypic plasticity is a
mechanism organisms may use to avoid habitat disturbances and threats. Previous plasticity studies of container-breeding
mosquitoes have examined the effect of habitat drying on the metamorphosis of Ochlerotatus triseriatus (Juliano et al, 1994), as well as the various effects
that environmental
influences may have on the hatch rate (Mogi, 1976; Edgerly et al, 1992;
Livdahl et al, 1984). We chose to examine the effects of cues that may
predict habitat risk where the cue itself is not a threat to the organism.
One such cue could be the frequency of hatching stimulation for a
container breeding mosquito. During plentiful rainfall conditions, eggs
are stimulated more frequently, while under possible drought conditions less frequent stimulation would
occur. Our previous work has indicated
that Aedes albopictus, the Asian Tiger Mosquito, does show a plastic response to the
frequency of hatch stimulation (Vitek
et al, 2003). Eggs that are stimulated
less frequently show less hatch delay than eggs that are stimulated more frequently. Here, we examine the hatch response of
another container breeding mosquito,
Ochlerotatus triseriatus. We tested a
variety of populations collected from areas in the northeastern USA that have a range of natural
frequencies of rainfall, as well differences in mean annual temperature and mean annual precipitation. We expected
that any exhibited plasticity should be directly related to the natural habitat.
Methods
In
the summer of 2002, eggs were collected from 7 locations in northeastern USA
(see figure 1). These sites were chosen based on
records of precipitation collected from National Weather Service stations (see
table 1). These data were collected
over the span of 15 to 54 years (depending on the site), and consisted of the number of days per month
with a minimum of .01 inches of precipitation.
Seven sites were chosen, including 4 sites at the upper end
of the distribution and 3 sites with relatively low rainfall frequency. Eggs
were collected after one month and brought into the laboratory. They were stored at a constant temperature
(23° C) and at a humidity of
90% or greater, in a 16:8 h light:dark cycle.
Half of the eggs were hatched and the larvae reared to
adulthood. These adults were force
mated to produce an F1 generation and stored under identical conditions as
the parental eggs. Experimental eggs
were drawn from both the wild-collected eggs and the F1 generation of
laboratory eggs. Eggs were divided into
batches of 50 and then subjected to one
of two levels of a stimulation frequency treatment (3 or 7 days between
stimuli). Hatch stimuli consisted of submerging the eggs into a
.5g/l nutrient broth mixture for 24 hours.
At the end of the stimulation period, hatched larvae were
removed and recorded and the eggs were removed and placed back into storage
until the next stimulation. This process
was repeated for 10 stimuli for all egg batches. At the conclusion of the experiment, unhatched eggs
were dissected to determined viability.
Non-viable eggs were removed from the analysis to determine an
accurate hatch percentage.
For all analyses
conducted, egg batches that had 100% hatch after the first stimulus was
excluded as the eggs were not exposed to differing treatments. The initial analysis tested the hatch rates
during the first stimulus to determine if there was any difference between the two
treatments. A secondary analysis was
conducted comparing
batches that had no subsequent eggs hatch to batches that had eggs hatch after
the first stimulus. A third analysis compared the hatch
rates for egg batches that did have any eggs hatching after the first
stimulus. For this analysis, a 50% critical
stimulus number was estimated using logistic regression. This number would be the hatch stimulus at which 50% or
more of the remaining eggs after the first stimulus had hatched. For all analyses, a variety of
environmental influences were tested including latitude, rainfall frequency,
mean annual precipitation, and mean annual temperature.
Results
The initial
analysis was to test for treatment effects and egg type (wild or force mated)
and population differences in hatch rate on the first stimulus (see figure
2). Ideally, treatment would have no
effect on the hatch rate at the first stimulus because the egg batches were not
subjected to different treatments at that time. There was no significant influence of
treatment on the hatch rates in the first stimulus (F = 2.7343, p =
.1025). Egg type (wild collected or laboratory
force mated) did have a significant influence (see table 2) with a p value of <.0001. Mean annual precipitation also had a
significant positive influence on the first hatch rate (see figure 3).
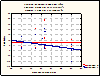
Figure 4. The interaction effect between treatment and rainfall frequency. More frequent rainfall results in a greater degree of plasticity between high and low frequency stimulation.
Table 1. Summary of
the sites
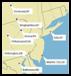
Figure 1. The seven
selected sites from which eggs were collected. Sites
were chosen based on the frequency of rainfall during the summer months, three sites with a low natural frequency
(indicated by a red marker) and four sites with a high natural frequency (indicated by a blue marker). The eighth site (Binghamton) was excluded from
this study.

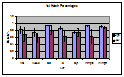
Figure 2. The hatch percentages of the first stimulus for each site and treatment (either high or low frequency of stimulation), with standard error bars.
Table 2. Summary of
ANOVA results testing the hatch rate during the first stimulus against egg type,
treatment, rainfall
frequency, and Mean Annual Precipitation.
Treatment is not significant, although both egg type (wild versus laboratory eggs) and Mean Annual Precipitation are significant.
Summary
These results
seem to indicate that hatch plasticity is used by Ochlerotatus triseriatus in response to stimulation
frequency. In addition, the plasticity
response does seem to vary as a result of the natural habitat. The two forms of plasticity analyzed
(hatching versus no hatching after the first hatch stimulus and the hatch rate
after the first stimulus) both show significant interactions between the treatment and
environmental factors including latitude, mean annual precipitation, and
rainfall frequency. This indicates that
plasticity responses may vary in the wild based on the natural conditions and that
indirect cues such as rainfall frequency may be used as predictors of habitat stability. One possible source of error in this experiment is small sample
size. Limited success with force mating
reduced the number of laboratory reared eggs available and forced the inclusion of
wild collected eggs. In addition a large number of the egg batches
had a 100% hatch rate on the first stimulus, limiting the number of replicates for each
treatment that were exposed to later stimuli.

Table 5. Summary of ANOVA results. Treatment interacted significantly with mean annual precipitation and rainfall frequency.
Literature Cited
Edgerly, J.S., Marvier, M.A.
1992. To Hatch of Not to
Hatch? Egg Hatch Response to Larval
Density and to Larval Contact in a Treehole Mosquito. Ecological Entomology, 17, 28-32.
Juliano, S.A., Stroffregen, T.L. 1994.
Effects of Habitat Drying on Size at and Time to Metamorphosis in the
Tree Hole Mosquito Aedes
triseriatus. Oecologia 97, 369-376.
Livdahl, T.P., Koenekoop, R.K., Futterweit, S.G. 1984.
The Complex Hatching Response of Aedes Eggs to Larval Density.
Ecological Entomology, 9, 437-442.
Mota, M. 1976. Variation in Oviposition, Hatching Rate, and
Setal Morphology in Laboratory Strains of Aedes Albopictus.
Mosquito News, 42, 196-201.
Vitek, C.J., Livdahl, T.P.
2003. Phenotypic Plasticity in
Hatch Rate and Development Rate of Aedes albopictus. American
Mosquito Control Association Annual Conference.
Minnesota, MN, USA. March 2003.
A copy of this poster can be found at http://posters.vitekweb.com/
The author can be contacted at cvitek@clarku.edu

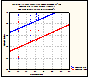
Figure
3. 1st hatch fraction graphed
against Mean
Annual Precipitation, with egg type indicated. There is no
interaction, but each influence is significant.
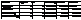
Egg batches were
then tested for a hatch response after the first stimulus. They were categorized as either “non responders” (replicates with viable
eggs remaining that failed to hatch) or “responders” (replicates with viable
egg remaining,
at least some of which did hatch). Egg
type (wild versus laboratory collected eggs) was tested initially, along with environmental factors,
to determine if conditions these eggs were exposed to in the wild resulted in
the eggs
not responding to subsequent hatch stimuli.
ANOVA results indicate that egg type did not play a role in the possible response or non response
of eggs to subsequent hatch stimulation (see table 3). A second analysis, incorporating the treatment factor,
was then conducted, and indicated that treatment interacted with latitude to produce a significant influence on
the egg’s response to subsequent hatch stimulation (the treatment*mean annual temperature interaction was almost
significant). Rainfall frequency was
also significant (see table 4), but this would not be a plasticity response
because there is no interaction with treatment.
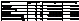

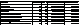
The 50% critical
stimulus number was estimated using a logistic regression analysis for each egg batch that had any hatch after
the first stimulus. This critical
stimulus number represents the estimated stimulus at which 50% of the remaining
eggs would hatch. Using this predicted stimulus, environmental factors and
treatment (high or low frequency stimulation) were analyzed with ANOVA. The environmental factors were mean annual
precipitation, mean annual temperature, latitude, and rainfall frequency. Treatment interacted significantly with both rainfall frequency and mean
annual precipitation (see table 5).
Figures 4 and 5 show this interaction effect, with regression lines drawn
for both of the two treatments.
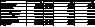
Table 3. ANOVA
results testing egg type as a factor for responsive or non responsive egg hatch after the 1st stimulus
Table 4. ANOVA
results testing the responders and non responders with treatment and environmental factors.
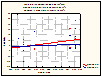

Figure 5. The interaction effect between treatment and mean annual precipitation. Greater precipitation results in a greater degree of plasticity between high and low frequency stimulation.