

Phenotypic plasticity in hatch rate and development rate of Aedes albopictus
Christopher J.
Vitek, Todd P. Livdahl

Biology Department,
Clark University, 950 Main St., Worcester, MA 01610
Abstract
Phenotypic plasticity is a well
studied phenomenon. Less studied, however, are plasticity responses to possible cues about random future
habitat disturbances. This study tested the effect of such cues on the hatch rate and development
rate of Aedes albopictus. Hatch stimulus timing was used as an indicator of habitat longevity
and probability of habitat desiccation.
Aedes albopictus eggs were hatched at one of two frequencies of
stimulation, representing
either a high frequency rainfall (less chance of habitat desiccation) or low frequency rainfall (greater chance of habitat
desiccation). The hatch rates and development rates between the two treatments were measured and
compared. The hatch rates of Aedes
albopictus that were subjected to low frequency stimulation
showed less hatch delay than the high
frequency stimulation treatment. The low frequency treatment also had a
faster mean development
time, although the difference was not statistically
significant. Additionally, the eggs that were hatched at a lower stimulation frequency
showed a greater sensitivity to larval density in regards to development time. The results
indicate that Aedes albopictus uses plasticity in response to frequency of stimulation, indicating that
they may be able to detect and respond to cues about random future habitat disturbances.
Methods
Eggs from a strain of Aedes albopictus originating near Kobe, Japan, eggs were collected within 24 hours after being laid by blood-fed females. These eggs were then subjected to one of two
experimental treatments – being
stimulated to hatch every 3 days or every 7 days (referred to as high frequency or frequent, and low frequency or
infrequent respectively). The hatch
stimulus consisted of submerging
the eggs for 24 hours in an aerated nutrient broth mixture. After 24 hours, the eggs were removed and stored in an incubator
at a minimum humidity of 90%, and minimum
temperature of 23 degrees C, with a cycle of 16 hours light/8 hours dark.
Larvae that hatched
were removed, counted, and then reared in a petri dish. The eggs were subjected to 10 sequential stimuli, after which the remaining unhatched
eggs were dissected to determine viability.
Hatched larvae were reared in
identical environments (with some slight variations in larval density). Habitat
size, food abundance, temperature, and light/dark cycles were identical for
all larvae. Upon pupation, larvae were removed from the
petri dishes and stored in individual vials until adult emergence.
The date of emergence was recorded, allowing for a total development time (from hatch to adulthood) to be recorded.
Analysis
of hatch rates
This
analysis focused on the hatch rates of eggs that did not hatch on the first
stimulus. Prior to the first stimulus, there
was no difference between the low and high frequency treatments, and no difference was observed
between hatch rates for the first stimulus (t=.789, p = .43). Using the viability data gathered
from the dissected eggs and removing any eggs that hatched during the first stimulus, the total
number of viable remaining eggs (after the first stimulus) was determined. Figure 1 shows the cumulative
hatch fractions of the remaining eggs for each stimulus after the first, with the high frequency or
low frequency treatment indicated. The number of stimuli
needed to reach a 50% hatch rate of the remaining eggs was calculated. Treatments that resulted in more hatch delay would take a
higher number of stimuli to reach the 50% threshold. A value of 11 stimuli was used for
experimental replicates that did not reach the 50% level by the 10th stimulus because that would be the
minimum number of stimuli needed to reach the 50% level. Using this method of analysis, there
was a significant difference between the two treatments – high frequency and low
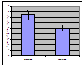
frequency stimulation (p = .0064) (Figure 2).
The low frequency treatment required an average of 5.05 stimuli to reach 50%
hatch, and the high frequency treatment required an average of 7.65
stimuli. The analysis results,
including the hatch rate for the first stimulus and the total number of
viable eggs for each replicate as additional factors, are summarized in Table
1. Neither of these two
variables had any significant influence on the time to reach the 50% threshold. This analysis indicates that the eggs
exposed to a low frequency stimulation undergo less hatch delay than
eggs exposed to a high frequency of stimulation.
Discussion
Many plasticity studies have looked at the effects of
habitat shrinking or drying on an aquatic organisms’ development, including
organisms such as frogs, salamanders, and container-breeding mosquitoes such
as Ochlerotatus triseriatus (Semlitsch, 1988; Crump, 1989;
Juliano et al., 1994). The cue provided in this study differed in a number of subtle
ways from those of other plasticity studies.
The actual hatch stimulus is an indicator of a suitable habitat (the submersion
in liquid indicates the habitat is ready for larvae), and the environmental cue
(a decrease in the frequency of stimulation) is an indicator of possible future habitat risk,
as opposed to an actual change in the organism’s habitat. There are a number of interpretations for the plasticity
responses observed. In terms of the
hatch rate,
it is possible that the mosquitoes are somehow “keeping track” of the frequency of stimulation. It is also possible that physical or
chemical changes in the actual chorion structure may result from being exposed
to air for varying
lengths of time (a possible maternal influence) and this change may influence the hatch rate. Larval development rate differences and
sensitivity to larval density may be in response to differences in the
frequency of stimulation, indicating that the mosquitoes may have been able
to detect environmental
cues and respond to them in ways other than simply by altering their hatch rates. If this is the case, these differences may
be the result
of different development trajectories brought on by the differences in frequency of stimulation. However, the development rate response is
more complex
than anticipated, and should be examined in more detail.
Figure 2. Comparison of
the number of stimuli needed to reach 50% hatch for the two treatments, high frequency
stimulation and low frequency stimulation, with error bars.
Table 1.
Summary of ANOVA results, testing the three variables. Stimulus frequency is the only significant influence, and
there are no significant interactions.
Figure 1. Cumulative
hatch fractions for the low frequency and high frequency treatments. If the hatch fraction reaches 1, then all of the eggs have
hatched. The eggs that hatched on the first stimulus are not
included. Nonviable eggs were removed
from the analysis.
Analysis
of development time
For larvae, the
time of development was calculated as the difference between the date of
hatching and
the date of emergence. These values
were calculated for larvae that hatched and survived to emergence. Data were recorded only for female
mosquitoes. In addition, the larval
density in the petri dish (ranging from 1 to 18) and the stimulus number
(during which the larvae hatched) were also recorded. For the analysis of the development rate, all larvae
that hatched in the first stimulus were ignored, for the same reason that the first
stimulus was ignored in the hatch analysis, i.e., at that time the two treatments were the same. To confirm the validity of this approach, an
analysis of the first stimulus hatchers shows that there was no
significant difference between the development time of the high and low
frequency treatments (t = .562, p = .5755).
For these first hatchers, larval density and stimulus
number were not included in the analysis (the larval density was 15 for each, and the stimulus number
was 1).
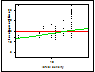
Figure 4.
Developmental time versus larval density of larvae exposed to either a high frequency
stimulation (red line) or low frequency stimulation (green line)
Table 2. Summary of
ANOVA results of the development time variables. As an individual factor, larval density was the only significant
variable. However, larval density interacted with
both stimulus frequency and stimulus number to produce a significant effect.
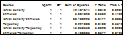

Analysis of
development time (cont.)
For the larvae
that hatched after the first stimulus, an ANOVA of development time in response to stimulus frequency
and larval density reveals a significant interaction between these factors
(Table 2, Fig. 4). The high frequency treatment showed no significant effect of
larval density on development time (p = .9790) while the low frequency treatment
did show a significant influence of larval density on the development rate of the
larvae (p=.0006). It appears that the
larvae that are hatched at a low frequency of stimulation are more
sensitive to larval density, having a slower development rate when there are more
larvae present.
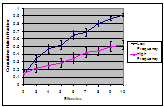


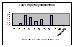
Figure 3. Frequency
histograms of the number of stimuli required to reach 50% hatch
Literature Cited
Crump, Martha L., 1989.
Effect of Habitat Drying on Development Time and Size at Metamorphosis
in Hyle pseudopuma.
Copeia 1989 (3): 794-797
Juliano, S. A., Stoffregen, T. L. 1994.
Effect of Habitat Drying on Size at and Time to Metamorphosis in the
Tree Hole Mosquito Aedes
triseriatus. Oecologia 97: 369-376
Semlitsch, R.D.
1988. Effects of Pond Drying
Time on the Metamorphosis and Survival in the Salamander Ambystoma talpoideum.
Copeia 1988 (4): 978 - 983
A copy of this poster can be found at
The author can be contacted at cvitek@clarku.edu